
Evaluation of IL-35, as a Possible Biomarker for Follow-Up after Therapy, in Chronic Human Schistosoma Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Measurement of Serum Cytokines
2.3. Statistical Analysis
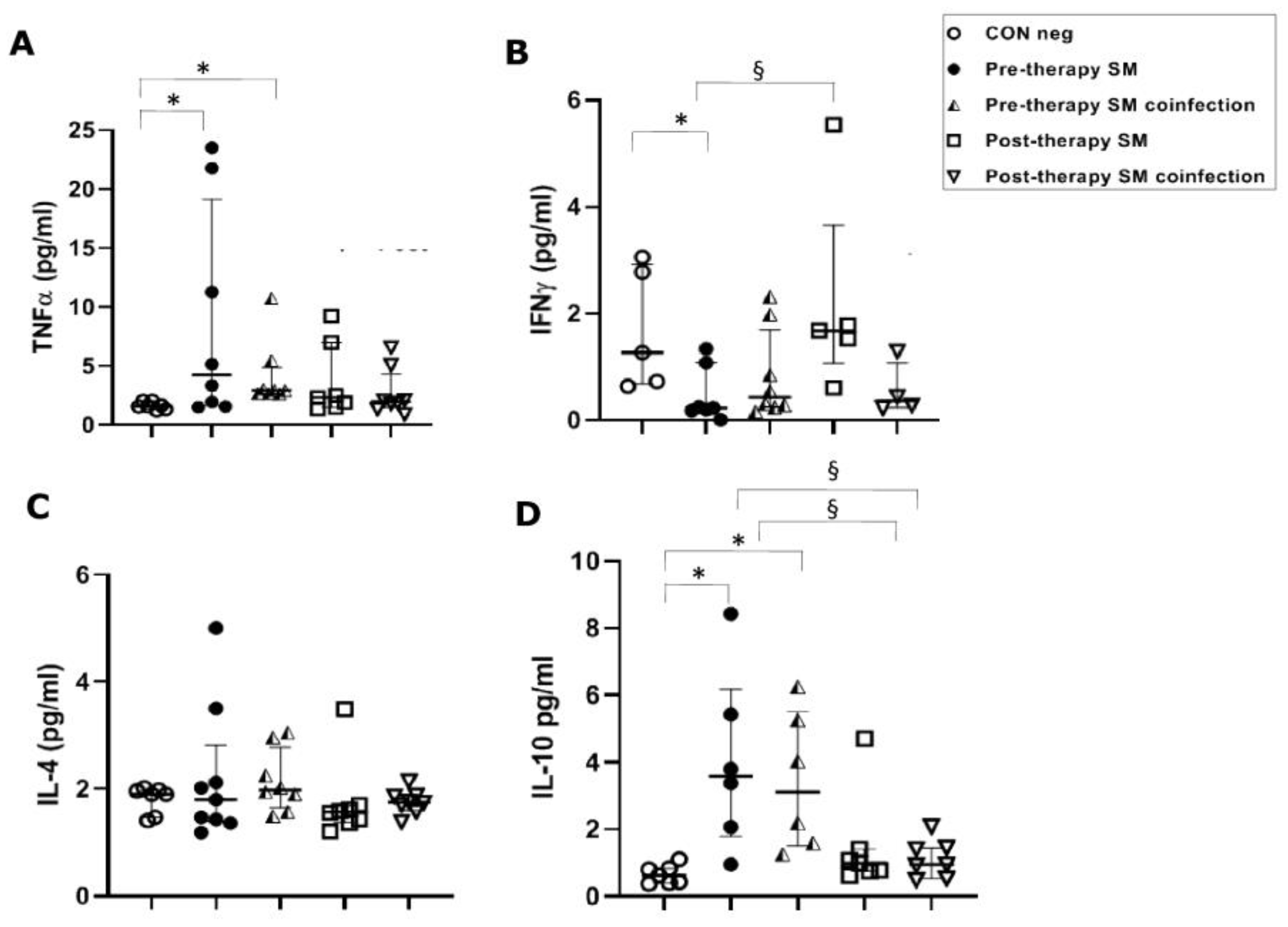
3. Results
3.1. Study Cohort
3.2. Th1 Response
3.2.1. TNFα Serical Levels before and after Anti-Parasitic Therapy
3.2.2. IFN-γ Serical Levels before and after Anti-Parasitic Therapy
3.3. Th2 Response
IL-4 Serical Levels before and after Anti-Parasitic Therapy
3.4. Treg Response
IL-10 Serical Levels before and after Anti-Parasitic Therapy
3.5. Breg Response
IL-35 Serical Levels before and after Anti-Parasitic Therapy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boon, N.A.M.; Fannes, W.; Rombouts, S.; Polman, K.; Volckaert, F.A.M.; Huyse, T. Detecting hybridization in African Schistosoma species: Does egg morphology complement molecular species identification? Parasitology 2017, 144, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Verjee, M.A. Schistosomiasis: Still a Cause of Significant Morbidity and Mortality. Res. Rep. Trop. Med. 2019, 10, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, C.; Rodríguez-Alonso, B.; López-Bernús, A.; Almeida, H.; Galindo-Pérez, I.; Velasco-Tirado, V.; Marcos, M.; Pardo-Lledías, J.; Belhassen-García, M. Clinical Spectrum of Schistosomiasis: An Update. J. Clin. Med. 2021, 10, 5521. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, M.; Torti, C.; Greco, G.; Strazzulla, A.; Costa, C.; Pisani, V.; Sorace, C.; Giancotti, A.; Lamberti, A.; Barreca, G.S.; et al. Prevalence of parasitic infections in migrants: Do official symptom-driven guidelines apply to the current situation? Infez. Med. 2018, 26, 347–355. [Google Scholar] [PubMed]
- Webster, B.L.; Diaw, O.T.; Seye, M.M.; Webster, J.P.; Rollinson, D. Introgressive hybridization of Schistosoma haematobium group species in Senegal: Species barriers break down between ruminant and human schistosomes. PLoS Negl. Trop. Dis. 2013, 7, e2110. [Google Scholar] [CrossRef]
- Sene-Wade, M.; Marchand, B.; Rollinson, D.; Webster, B.L. Urogenital schistosomiasis and hybridization between Schistosoma haematobium and Schistosoma bovis in adults living in Richard-Toll. Senegal. Parasitol. 2018, 145, 1723–1726. [Google Scholar] [CrossRef]
- Marascio, N.; Loria, M.T.; Lamberti, A.G.; Pavia, G.; Adams, N.J.; Quirino, A.; Divenuto, F.; Mazzitelli, M.; Greco, G.; Trecarichi, E.M.; et al. Molecular characterization of Schistosoma infections in African migrants: Identification of a Schistosoma haematobium-bovis hybrid in bladder biopsies. J. Travel Med. 2022, 29, taab194. [Google Scholar] [CrossRef]
- White, M.P.J.; McManus, C.M.; Maizels, R.M. Regulatory T-cells in helminth infection: Induction, function and therapeutic potential. Immunology 2020, 160, 248–260. [Google Scholar] [CrossRef]
- Khan, A.R.; Amu, S.; Saunders, S.P.; Fallon, P.G. The generation of regulatory B cells by helminth parasites. Methods Mol. Biol. 2014, 1190, 143–162. [Google Scholar]
- Maizels, R.M. Regulation of Immunity and allergy by helminth parasites. Allergy 2020, 75, 524–534. [Google Scholar] [CrossRef]
- Paixão de Souza, R.; Araújo, M.I.; Lopes, D.M.; Oliveira, S.C.; Fernandes, J.S.; de Jesus, K.E.M.; Carvalho, E.M.; Oliveira, R.R.; Cardoso, L.S. Profile of T and B lymphocytes in individuals resistant to Schistosoma mansoni infection. Parasitol. Res. 2022, 121, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, C.T.; Sanin, D.E.; Cook, P.C.; Mountford, A.P. CD4+ T cell hyporesponsiveness after repeated exposure to Schistosoma mansoni larvae is dependent upon interleukin-10. Infect. Immun. 2015, 83, 1418–1430. [Google Scholar] [CrossRef] [PubMed]
- Castro, V.N.; Rodrigues, J.L.; Cardoso, D.T.; Resende, S.D.; Magalhães, F.C.; Souza, D.C.; Requeijo, M.H.; Negrão-Corrêa, D.; Geiger, S.M. Systemic Cytokine and Chemokine Profiles in Individuals With Schistosoma mansoni Infection and Low Parasite Burden. Front. Immunol. 2018, 9, 2975. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.M.; Fathy, G.M.; Abdel-Rahman, S.A.; El-Shafei, M.A. Cytokine patterns in experimental Schistosoma mansoni infected mice treated with silymarin. J. Parasit. Dis. 2016, 40, 922–929. [Google Scholar] [CrossRef]
- Schmiedel, Y.; Mombo-Ngoma, G.; Labuda, L.A.; Janse, J.J.; de Gier, B.; Adegnika, A.A.; Issifou, S.; Kremsner, P.G.; Smits, H.H.; Yazdanbakhsh, M. CD4+CD25hiFOXP3+ Regulatory T Cells and Cytokine Responses in Human Schistosomiasis before and after Treatment with Praziquantel. PLoS Negl. Trop. Dis. 2015, 9, e0003995. [Google Scholar] [CrossRef]
- Tweyongyere, R.; Nassanga, B.R.; Muhwezi, A.; Odongo, M.; Lule, S.A.; Nsubuga, R.N.; Webb, E.L.; Cose, S.C.; Elliott, A.M. Effect of Schistosoma mansoni infection and its treatment on antibody responses to measles catch-up immunisation in pre-school children: A randomised trial. PLoS Negl. Trop. Dis. 2019, 13, e0007157. [Google Scholar] [CrossRef]
- Rocamora-Reverte, L.; Melzer, F.L.; Würzner, R.; Weinberger, B. The Complex Role of Regulatory T Cells in Immunity and Aging. Front. Immunol. 2021, 11, 616949. [Google Scholar] [CrossRef]
- Xiao, J.; Guan, F.; Sun, L.; Zhang, Y.; Zhang, X.; Lu, S.; Liu, W. B cells induced by Schistosoma japonicum infection display diverse regulatory phenotypes and modulate CD4+ T cell response. Parasites Vectors 2020, 13, 147. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Jiang, J.; She, X.; Niu, Y.; Ming, Y. The Potential Role of Schistosome-Associated Factors as Therapeutic Modulators of the Immune System. Infect. Immun. 2020, 88, e00754-19. [Google Scholar] [CrossRef]
- Costain, A.H.; Phythian-Adams, A.T.; Colombo, S.A.P.; Marley, A.K.; Owusu, C.; Cook, P.C.; Brown, S.L.; Webb, L.M.; Lundie, R.J.; Smits, H.H.; et al. Dynamics of Host Immune Response Development During Schistosoma mansoni Infection. Front Immunol. 2022, 13, 906338. [Google Scholar] [CrossRef]
- Ogongo, P.; Nyakundi, R.K.; Chege, G.K.; Ochola, L. The Road to Elimination: Current State of Schistosomiasis Research and Progress Towards the End Game. Front. Immunol. 2022, 13, 846108. [Google Scholar] [CrossRef] [PubMed]
- Masamba, P.; Kappo, A.P. Immunological and Biochemical Interplay between Cytokines, Oxidative Stress and Schistosomiasis. Int. J. Mol. Sci. 2021, 22, 7216. [Google Scholar] [CrossRef]
- Taylor, M.D.; van der Werf, N.; Maizels, R.M. T cells in helminth infection: The regulators and the regulated. Trends Immunol. 2012, 33, 181–189. [Google Scholar] [CrossRef]
- Labuda, L.A.; Adegnika, A.A.; Rosa, B.A.; Martin, J.; Ateba-Ngoa, U.; Amoah, A.S.; Lima, H.M.; Meurs, L.; Mbow, M.; Manurung, M.D.; et al. A Praziquantel Treatment Study of Immune and Transcriptome Profiles in Schistosoma haematobium-Infected Gabonese Schoolchildren. J. Infect. Dis. 2020, 222, 2103–2113. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Roch, T.; Lampropoulou, V.; O’Connor, R.A.; Stervbo, U.; Hilgenberg, E.; Ries, S.; Dang, V.D.; Jaimes, Y.; Daridon, C.; et al. IL-35-producing B cells are critical regulators of immunity during autoimmune and infectious diseases. Nature 2014, 507, 366–370. [Google Scholar] [CrossRef]
- Ye, C.; Yano, H.; Workman, C.J.; Vignali, D.A.A. Interleukin-35: Structure, Function and Its Impact on Immune-Related Diseases. J. Interferon Cytokine Res. 2021, 41, 391–406. [Google Scholar] [CrossRef]
- Haeberlein, S.; Obieglo, K.; Ozir-Fazalalikhan, A.; Chayé, M.A.M.; Veninga, H.; van der Vlugt, L.E.P.M.; Voskamp, A.; Boon, L.; den Haan, J.M.M.; Westerhof, L.B.; et al. Schistosome egg antigens, including the glycoprotein IPSE/alpha-1, trigger the development of regulatory B cells. PLoS Pathog. 2017, 13, e1006539. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, M.; Matera, G.; Votino, C.; Visconti, F.; Strazzulla, A.; Loria, M.T.; Peronace, C.; Settembre, P.; Giancotti, A.; Liberto, M.C.; et al. A case report of Schistosoma haematobium infection in a pregnant migrant raises concerns about lack of screening policies. J. Travel Med. 2016, 24, taw076. [Google Scholar] [CrossRef]
- Cucchetto, G.; Buonfrate, D.; Marchese, V.; Rodari, P.; Ferrari, A.; Zanotti, P.; Bottieau, E.; Silva, R.; Bisoffi, Z.; Gobbi, F. High-dose or multi-day praziquantel for imported schistosomiasis? A systematic review. J Travel Med. 2019, 26, taz050. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC) Guidelines 2020. Resources for Health Professionals–Parasites-Schistosomiasis. Available online: https://www.cdc.gov/parasites/schistosomiasis/health_professionals/index.html#tx (accessed on 9 April 2023).
- Silva, P.C.V.; Gomes, A.V.; de Britto, L.R.P.B.; de Lima, E.L.S.; da Silva, J.L.; Montenegro, S.M.L.; Muniz, M.T.C.; Domingues, A.L.C. Influence of a TNF-α Polymorphism on the Severity of Schistosomiasis Periportal Fibrosis in the Northeast of Brazil. Genet. Test. Mol. Biomark. 2017, 21, 658–662. [Google Scholar] [CrossRef]
- Mutengo, M.M.; Mduluza, T.; Kelly, P.; Mwansa, J.C.L.; Kwenda, G.; Musonda, P.; Chipeta, J. Low IL-6, IL-10, and TNF-α and High IL-13 Cytokine Levels Are Associated with Severe Hepatic Fibrosis in Schistosoma mansoni Chronically Exposed Individuals. J. Parasitol. Res. 2018, 2018, 9754060. [Google Scholar] [CrossRef] [PubMed]
- Egesa, M.; Lubyayi, L.; Tukahebwa, E.M.; Bagaya, B.S.; Chalmers, I.W.; Wilson, S.; Hokke, C.H.; Hoffmann, K.F.; Dunne, D.W.; Yazdanbakhsh, M.; et al. Schistosoma mansoni schistosomula antigens induce Th1/Pro-inflammatory cytokine responses. Parasite Immunol. 2018, 40, e12592. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Zhang, J.; Chen, H.; Nie, H.; Miller, H.; Gong, Q.; Liu, C. T Lymphocyte-Mediated Liver Immunopathology of Schistosomiasis. Front. Immunol. 2020, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.S.; Mentink-Kane, M.M.; Pesce, J.T.; Ramalingam, T.R.; Thompson, R.; Wynn, T.A. Immunopathology of Schistosomiasis. Immunol. Cell Biol. 2007, 85, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Divenuto, F.; Pavia, G.; Marascio, N.; Barreca, G.S.; Quirino, A.; Matera, G. Role of Treg, Breg and other cytokine sets in host protection and immunopathology during human leishmaniasis: Are they potential valuable markers in clinical settings and vaccine evaluation? Acta Trop. 2023, 240, 106849. [Google Scholar] [CrossRef] [PubMed]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef]
- Fattori, A.C.M.; Montija, E.A.; Fragelli, B.D.L.; Correia, R.O.; de Castro, C.A.; Romanello, L.; Nogueira, C.T.; Allegretti, S.M.; Soares, E.G.; Pereira, H.D.; et al. Effects of Immunization with Recombinant Schistosoma mansoni Enzymes AK and HGPRT: Murine Infection Control. Pathogens 2023, 12, 69. [Google Scholar] [CrossRef]
- Meningher, T.; Barsheshet, Y.; Ofir-Birin, Y.; Gold, D.; Brant, B.; Dekel, E.; Sidi, Y.; Schwartz, E.; Regev-Rudzki, N.; Avni, O.; et al. Schistosomal extracellular vesicle-enclosed miRNAs modulate host Thelper differentiation. EMBO Rep. 2020, 21, e47882. [Google Scholar] [CrossRef]
- Ondigo, B.N.; Ndombi, E.M.; Nicholson, S.C.; Oguso, J.K.; Carter, J.M.; Kittur, N.; Secor, W.E.; Karanja, D.M.S.; Colley, D.G. Functional Studies of T Regulatory Lymphocytes in Human Schistosomiasis in Western Kenya. Am. J. Trop. Med. Hyg. 2018, 98, 1770–1781. [Google Scholar] [CrossRef]
- Nogueira, R.A.; Lira, M.G.S.; Licá, I.C.L.; Frazão, G.C.C.G.; Dos Santos, V.A.F.; Filho, A.C.C.M.; Rodrigues, J.G.M.; Miranda, G.S.; Carvalho, R.C.; Nascimento, F.R.F. Praziquantel: An update on the mechanism of its action against schistosomiasis and new therapeutic perspectives. Mol. Biochem. Parasitol. 2022, 252, 111531. [Google Scholar] [CrossRef]
- Eyoh, E.; McCallum, P.; Killick, J.; Amanfo, S.; Mutapi, F.; Astier, A.L. The anthelmintic drug praziquantel promotes human Tr1 differentiation. Immunol Cell Biol. 2019, 97, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, J.I.; Hsieh, M.H. Immune responses to Schistosoma haematobium infection. Parasite Immunol. 2014, 36, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Matera, G.; Loria, M.T.; Peronace, C.; Catanzariti, T.; Settembre, P.; Giancotti, A.; Lamberti, A.G.; Barreca, G.S.; Galati, L.; Dodaro, G.; et al. Increase of Vascular Endothelial Growth Factor and Decrease of MCP-1 and Some Updated Epidemiology Aspects of Cystic Echinococcosis Human Cases in Calabria Region. Mediat. Inflamm. 2018, 14, 4283672. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; Holland, M.J. Parasite Immunity: Pathways for expelling intestinal helminths. Curr. Biol. 1998, 8, R711–R714. [Google Scholar] [CrossRef]
- Marinho, F.V.; Alves, C.C.; de Souza, S.C.; da Silva, C.M.; Cassali, G.D.; Oliveira, S.C.; Pacifico, L.G.; Fonseca, C.T. Schistosoma mansoni Tegument (Smteg) Induces IL-10 and Modulates Experimental Airway Inflammation. PLoS ONE 2016, 11, e0160118. [Google Scholar] [CrossRef]
- Matera, G.; Giancotti, A.; Scalise, S.; Pulicari, M.C.; Maselli, R.; Piizzi, C.; Pelaia, G.; Tancrè, V.; Muto, V.; Doldo, P.; et al. Ascaris lumbricoides-induced suppression of total and specific IgE responses in atopic subjects is interleukin 10-independent and associated with an increase of CD25+ cells. Diagn. Microbiol. Infect. Dis. 2008, 62, 280–286. [Google Scholar] [CrossRef]
- Correale, J.; Marrodan, M.; CarneroContentti, E. Interleukin-35 is a critical regulator of immunity during helminth infections associated with multiple sclerosis. Immunology 2021, 164, 569–586. [Google Scholar] [CrossRef]
- Beltrame, A.; Guerriero, M.; Angheben, A.; Gobbi, F.; Requena-Mendez, A.; Zammarchi, L.; Formenti, F.; Perandin, F.; Buonfrate, D.; Bisoffi, Z. Accuracy of parasitological and immunological tests for the screening of human schistosomiasis in immigrants and refugees from African countries: An approach with Latent Class Analysis. PLoS Negl. Trop. Dis. 2017, 11, e0005593. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| ID Patient | Age | Sex | Country of Origin | Sample Collected | Parasite Identified by Microscopy |
|---|---|---|---|---|---|
| 1 | 23 | M | Guinea | Urine | S. haematobium |
| 2 | 21 | M | Sierra Leone | Urine | S. haematobium |
| 3 | 23 | M | Ivory Coast | Urine | S. haematobium |
| 4 | 19 | M | Ghana | Urine | S. haematobium |
| 5 | 23 | M | Mali | Urine | S. haematobium |
| 6 | 20 | M | Cameroon | Urine | S. haematobium |
| 7 | 20 | M | Liberia | Urine | S. haematobium |
| 8 | 20 | M | Brazil | Urine | S. haematobium |
| 9 | 29 | M | Ghana | Urine | S. haematobium |
| 10 | 32 | M | Mali | Urine | S. haematobium |
| 11 | 20 | F | Mali | Urine | S. haematobium |
| 12 | 21 | F | Mali | Urine | S. haematobium |
| 13 | 20 | M | Senegal | Urine | S. haematobium |
| 14 | 21 | M | Senegal | Urine | S. haematobium |
| 15 | 18 | M | Mali | Urine | S. haematobium |
| 16 | 18 | M | Guinea | Stool | S.mansoni |
| 17 | 22 | M | Senegal | Stool | S.mansoni |
| 18 | 19 | M | Senegal | Stool | S.mansoni |
| 19 | 29 | F | Ivory Coast | Stool | S.mansoni |
| 20 | 29 | M | Camerun | Stool | S.mansoni |
| 21 | 19 | M | Liberia | Stool | S.mansoni |
| 22 | 31 | M | Ghana | Stool | S.mansoni |
| 23 | 23 | M | Ivory Coast | Stool | S.mansoni |
| 24 | 25 | M | Gambia | Stool | S. mansoni |
| 25 | 22 | M | Bangladesh | Stool | S. mansoni |
| 26 | 21 | M | Ivory Coast | Stool | S. mansoni |
| 27 | 19 | M | Sierra Leone | Stool | S. mansoni, E.coli |
| 28 | 23 | M | Ivory Coast | Stool | S. mansoni, E.nana |
| 29 | 28 | M | Guinea | Stool | S. mansoni, Blastocystis spp. |
| 30 | 28 | M | Ethiopia | Stool | S. mansoni, Blastocystis spp. |
| 31 | 19 | M | Guinea | Stool | S. mansoni, G.intestinalis |
| 32 | 30 | M | Ivory Coast | Stool | S. mansoni, E. nana |
| 33 | 24 | F | Brazil | Stool | S. mansoni, E.nana, E. coli, C.mesnili |
| 34 | 19 | M | Mali | Stool | S. mansoni,I.butschlii,E.col, E. h/d/m |
| 35 | 33 | M | Mali | Bladder biopsy * | HybridS. haematobium-bovis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marascio, N.; Loria, M.T.; Pavia, G.; Peronace, C.; Adams, N.J.; Campolo, M.; Divenuto, F.; Lamberti, A.G.; Giancotti, A.; Barreca, G.S.; et al. Evaluation of IL-35, as a Possible Biomarker for Follow-Up after Therapy, in Chronic Human Schistosoma Infection. Vaccines 2023, 11, 995. https://doi.org/10.3390/vaccines11050995
Marascio N, Loria MT, Pavia G, Peronace C, Adams NJ, Campolo M, Divenuto F, Lamberti AG, Giancotti A, Barreca GS, et al. Evaluation of IL-35, as a Possible Biomarker for Follow-Up after Therapy, in Chronic Human Schistosoma Infection. Vaccines. 2023; 11(5):995. https://doi.org/10.3390/vaccines11050995
Chicago/Turabian StyleMarascio, Nadia, Maria Teresa Loria, Grazia Pavia, Cinzia Peronace, Neill James Adams, Morena Campolo, Francesca Divenuto, Angelo Giuseppe Lamberti, Aida Giancotti, Giorgio Settimo Barreca, and et al. 2023. "Evaluation of IL-35, as a Possible Biomarker for Follow-Up after Therapy, in Chronic Human Schistosoma Infection" Vaccines 11, no. 5: 995. https://doi.org/10.3390/vaccines11050995